Brian Potter's latest essay reframes sexual reproduction not as a biological curiosity but as a search algorithm, one that mirrors the modular tricks engineers use to build complex technology. The deeper claim is that evolution and engineering, separated by billions of years and operating in entirely different domains, stumbled onto the same insight about how to acquire information efficiently.
There is a particular kind of intellectual pleasure in watching two unrelated systems turn out to be solving the same problem. Brian Potter's essay on biological evolution and information acquisition delivers exactly that satisfaction, arguing that sexual reproduction and modular engineering are, at bottom, the same trick deployed in different substrates. The thesis is deceptively simple: both biology and technology face an enormous search problem, and both have discovered that the way to win is to break the problem into independently testable pieces. What makes the argument worth sitting with is not the novelty of any single claim but the way it dissolves the boundary we usually maintain between the designed and the evolved.
The Search Problem, Stated Plainly
Potter builds his case on a foundation he laid in an earlier piece about economist Brian Arthur's simulation of technological evolution, in which complex circuits like a 4-bit adder emerged from the random recombination of simple logic gates. The reason that simulation works, rather than wandering forever through a combinatorial wilderness, is modularity. Once the system discovers a working 3-way AND gate, it can lock that design in and treat it as a single building block, using it to construct a 4-way gate, and so on up the ladder of complexity. The search never has to guess an entire 68-gate circuit at once. It guesses one gate at a time and checks its work.
The combination lock analogy Potter uses is the clearest way into this. A five-digit lock with a hundred values per digit has ten billion possible combinations, and trying them blindly is hopeless. But a skilled lockpick who can hear a click when each individual digit lands correctly faces only five separate searches over a hundred values each, five hundred possibilities total rather than ten billion. The difference is not incremental. It is the difference between a tractable problem and an impossible one, and it comes entirely from being able to evaluate each part of the solution independently.
The genuinely interesting move in the essay is the claim that biological evolution found the same shortcut, and that it did so through something we rarely think of in computational terms: sex.
Why Asexual Reproduction Hits a Wall
To make the comparison concrete, Potter runs a stripped-down simulation. A population of 100 creatures, each carrying a genome of 200 genes, where every gene is either a 1 (good) or a 0 (bad). Fitness is just the count of good genes. Under asexual reproduction, each creature produces noisy copies of itself, with a small per-gene chance of mutation, and the fittest children survive to the next round. Fitness climbs, as you would expect, but it climbs into a headwind.
The headwind is worth understanding because it reveals something counterintuitive about pure mutation as a source of variation. Once a genome is mostly 1s, a random flip is more likely to ruin a good gene than to fix a bad one, simply because there are more good genes available to break. So for any above-average parent, the average child is worse. Evolution still makes progress because selection culls the losers and a lucky few children land higher than their parents, but the whole process is dragging a weight uphill. As the population approaches the maximum, the rate of improvement crawls toward flipping a single beneficial gene every four generations.
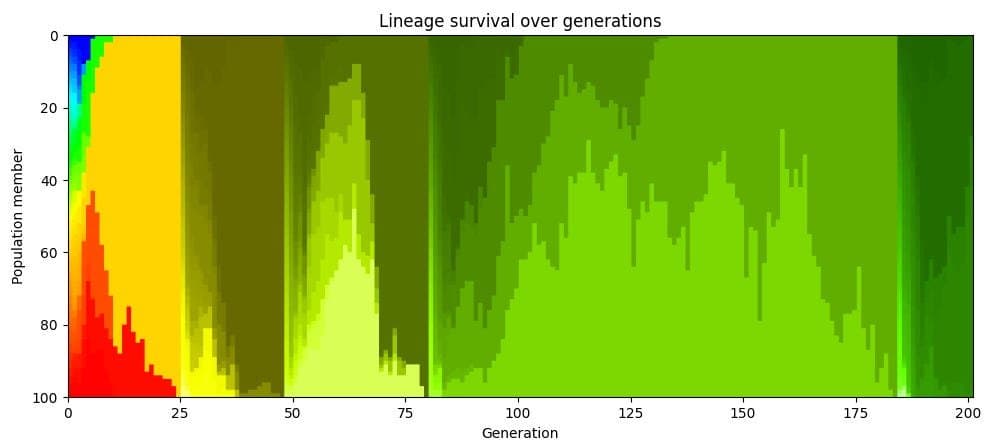
The deeper pathology is what Potter, following the evolutionary biology literature, calls clonal interference. In an asexual lineage, a beneficial mutation can only spread if the entire lineage carrying it outcompetes and eliminates every other lineage. The simulation makes this visible by color-coding each founding creature's descendants: within a few generations the whole population collapses to a single color, meaning everyone descends from one lucky ancestor. Every other lineage, along with whatever useful mutations it happened to discover, is erased. Two good ideas arising in two different individuals cannot be combined. They fight, and one of them dies.
Recombination as Parallel Search
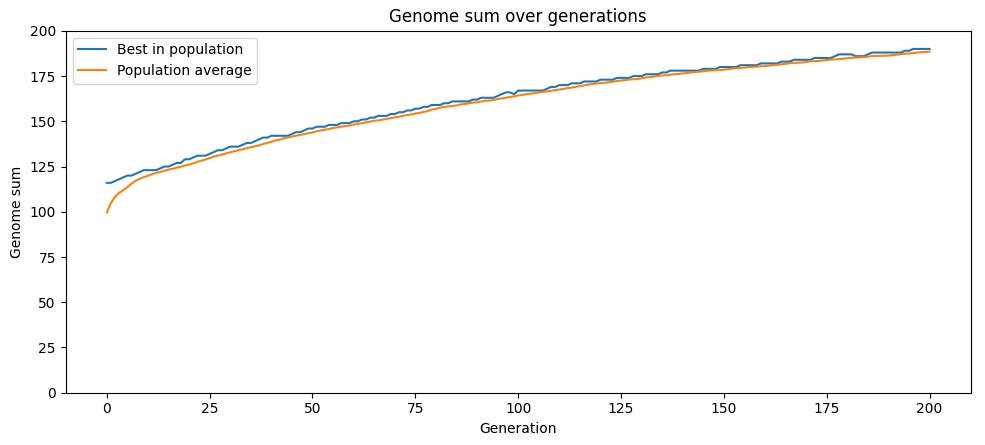
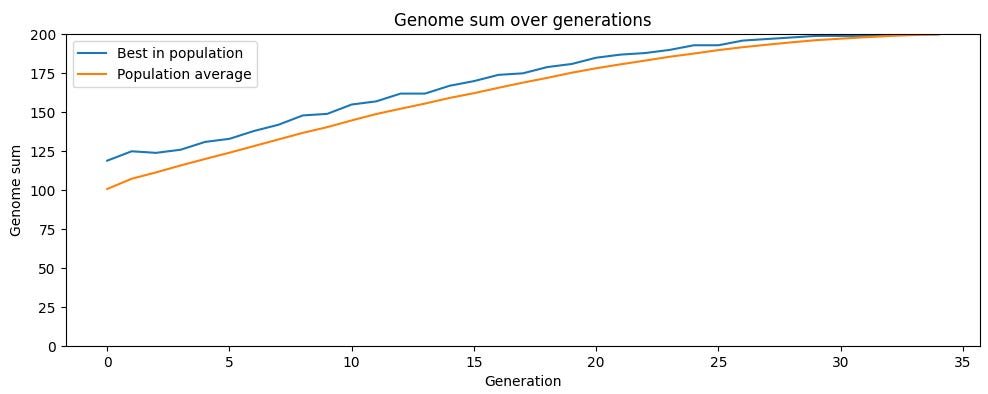
Sexual reproduction breaks this logjam, and the simulation results are dramatic. With no mutation at all, drawing each child's genes randomly from two parents, the population reaches maximum fitness in 33 generations. The asexual version was still stuck around 187 out of 200 after two hundred generations. The mechanism is that recombination introduces variation without degrading average fitness. A child who is a random shuffle of two parents has, on average, the same fitness as those parents, so selection operates on a distribution centered much higher than in the mutation-driven case.
Potter's framing is that recombination converts a single search over the best 200-gene genome into roughly 200 parallel searches for the best gene at each position. Each gene's value gets tested across many genetic backgrounds, and the good versions accumulate because they keep ending up in winning combinations. Returning to the lock, asexual reproduction with its single-click feedback is one kind of modular search, but sexual reproduction is subtler still: it is like trying many full combinations, receiving a score for how close each one is, and inferring which individual dials are correct from the pattern of scores. The information about each position is extracted in parallel rather than serially.
This is where the lineage mathematics becomes almost poetic. An asexual creature has one parent, one grandparent, one great-grandparent, a thread reaching back through time. A sexual creature has two parents, four grandparents, eight great-grandparents, a widening fan that lets beneficial variants from many ancestors converge into a single descendant. In the sexual simulation, genes from nearly 75% of the original population survive after 34 generations. In the asexual one, the figure is 1%, and shrinks further in larger populations. Sex is, among other things, a refusal to throw away good ideas just because they were born in the wrong body.
The Information-Theoretic Payoff
The synthesis Potter reaches for is to measure all of this in bits. A bit, in his gloss, is simply something that halves the number of possibilities you still have to consider. A freshly randomized population, split fifty-fifty on every gene, carries zero bits of information per gene because you are maximally uncertain about the right value. A population fixed on the correct value for every gene carries one bit per gene, full certainty. Total information acquired tracks fitness, and the sexual population accumulates it far faster than the asexual one.
This lets evolution and engineering be placed on the same axis. Arthur's nonmodular circuit search gleaned less than a millionth of a bit per attempt, a punishing crawl, while the modular gate-by-gate search pulled in roughly 0.003 bits per attempt, thousands of times faster. Biology's recombination is doing the analogous thing: it raises the bits-per-generation harvested from the genetic variation already present in the population. The thesis lands here. Modularity, in silicon or in DNA, is fundamentally a method for acquiring information about a vast search space without paying to examine the whole thing.
Where the Model Bends
Potter is honest about the load-bearing assumption, and it is a large one. The whole tidy picture depends on genes contributing to fitness independently, so that the value of gene 27 has nothing to do with gene 145. Real genomes are not like this. Genes interact, and the usefulness of a variant often depends entirely on which other variants accompany it. When fitness becomes coupled in this way, the search landscape grows rugged, full of peaks and valleys, and the clean parallel-search story gets complicated. He points to Stuart Kauffman's work on NK landscapes as the more rigorous treatment, and the gesture is the right one: the additive model is a teaching instrument, not a description of life.
The other honest caveat is that the asexual-versus-sexual dichotomy is too clean. Bacteria, the supposed exemplars of clonal reproduction, cheat constantly through horizontal gene transfer, passing genes laterally between existing organisms rather than only vertically to offspring. This is the primary route by which antibiotic resistance propagates, and by some estimates 20 to 80% of bacterial genomes are stitched together from transferred material. The lesson is not that sex is special but that recombination in any form, the ability to combine good genes discovered separately, is the prize. Life keeps inventing ways to get it.
What the Convergence Suggests
The reason this essay rewards reflection beyond its simulations is the structural claim it implies about how complexity gets built anywhere. Two processes that share no mechanism, no designer, and no common history converged on the same solution to the same abstract problem. That convergence hints that modularity and recombination are not contingent tricks but something closer to necessary features of any system that has to find rare good configurations in an astronomically large space within finite time. The space of working 68-gate adders is vanishingly small against 2^853 possibilities, and the space of viable genomes is similarly sparse. Brute force is not an option for evolution any more than it is for an engineer, and both arrived at the same answer because, perhaps, it is the only answer available.
There is a tension worth holding onto, though. Potter is careful to note that human technology is not actually built by random recombination; Arthur's randomness was a measuring instrument, a way to gauge how hard a search is under different conditions, not a model of how engineers think. Biology genuinely is random in its variation, with selection supplying the only direction. So the convergence is real but asymmetric. Evolution has no foresight and compensates with an enormous parallel search refined over deep time, while engineering smuggles in intention and can shortcut the search with understanding. Reading the two as instances of one information-acquisition principle illuminates both, yet it also quietly raises the question of how much of what we call design is just a faster, foresighted version of the same blind harvesting that built the whale and the bacterium.
For readers who want the rigorous version rather than the intuition pump, Potter points to chapter 19 of David MacKay's Information Theory, Inference, and Learning Algorithms, which is freely available and remains one of the better places to watch evolution and information theory be spoken about in the same breath. The essay itself lives at Construction Physics, where Potter usually writes about buildings and industrial technology, which makes this excursion into population genetics a small piece of evidence for his own thesis: good ideas travel best when they can be recombined across the lineages that first discovered them.
Comments
Please log in or register to join the discussion